TESSUTI TEGUMENTALI PRIMARI E SECONDARI
Introduzione
Rivestono tutta la pianta ed hanno funzione di protezione. I tessuti
tegumentali che rivestono le parti aeree hanno, come funzione principale
quella di impedire una traspirazione troppo elevata, che provocherebbe
l'avvizzimento della pianta, perciò sono costituiti, per la maggior
parte, da cellule a stretto contatto tra loro, senza spazi
intercellulari, e hanno le pareti impregnate da polimeri impermeabili:
la cutina o la suberina. Il tessuto tegumentale che riveste le parti non
legnose della pianta (fusti giovani, foglie, frutti, fiori) è l'epidermide, mentre le parti lignificate sono rivestite dal sughero.
Anche la radice è rivestita, nelle parti non lignificate, da un'epidermide, il rizoderma, che ha una funzione completamente diversa: quella di assorbire acqua e sali minerali dal terreno: anche qui le cellule sono strettamente a contatto tra di loro ma hanno parete molto permeabile e alcune cellule possiedono delle lunghe estroflessioni, i peli radicali, che servono per aumentarne la superficie assorbente.
Anche la radice è rivestita, nelle parti non lignificate, da un'epidermide, il rizoderma, che ha una funzione completamente diversa: quella di assorbire acqua e sali minerali dal terreno: anche qui le cellule sono strettamente a contatto tra di loro ma hanno parete molto permeabile e alcune cellule possiedono delle lunghe estroflessioni, i peli radicali, che servono per aumentarne la superficie assorbente.
Tessuti Tegumentali Primari----------->Tessuti Tegumentali esterni------->Epidermide - Esoderma
Tessuti Tegumentali primari---------------->tessuti tegumentali Interni-------> Endoderma
Epidermide: l'epidermide è tegumentale esterno di origine primaria. Ricopre l'intero corpo della pianta; esso deriva strado più superficiale dei meristemi apicali.
MERISTEMI APICALI-----> (PROTODERMA) -----> EPIDERMITE
EPIDERMIDE NELLE RADICI= RIZODERMA(CON FUNZIONI OPPOSTE ALL'EPIDERMIDE)
L'epidermide normalmente è monostratificata, sopratutto per le piante dei nostri climi; ma può essere pluristratificato nelle piante adatte a climi aridi (xerofite) come l'oleandro (Nerium oleander) o il ficus (Ficus elastica).


Funzione epidermide. La funzione dell'epidermide negli organi aerei della pianta è essenzialmente di protezione, grazie alle sue caratteristiche questo tessuto modera efficacemente la traspirazone, regola gli scambi gassosi, da una moderata protezione meccanica ed una buona difesa contro i parassiti. Per svolgere tali funzoni, l'epidermide è priva di spazi intercellulari, perchè attraverso questi spazi potrebbero penetrare organismi patogeni.
Funzioni accessorie dell'epidermide.
- Fotosintesi;
- Attività secretoria;
- Riserva d'acqua e dei prodotti del metabolismo;
- Meccanismi di movimento;
- Da origine ai meristemi da cui derivano gli apparati stomatici;
- Partecipa alla cicatrizzazione delle ferite;
- nelle piante EPIFITE , (piante che vivono su altre pante, felci, muschi, licheni) può funzionare come tessuto assorbente.
Struttura del tessuto epidermico
E' un tessuto polimorfo. Oltre a variare tra pianta e pianta, puo variare anche nella stessa pianta nei diversi organi.
Il tessuto è formato da diversi tipologie cellulari, in quanto in esso sono presenti:
- cellule epidermiche;
- Cellule di guardia degli stomi;
- Cellule specializzate come quelle dei tricomi.
LA CELLULA EPIDERMICA NEL TESSUTO E' PRIVA DI SPAZI INTERCELLULARI.
La MORFOLOGIA della cellula epidermica dipende dalla morfologia dell'organo che va a ricoprire.
Le cellule epidermiche sono vive, il loro citoplasma è di solito ridotto ad un sottile strato parietale.Nel grande vacuolo centrale può essere disciolti pigmenti, che consentono all'epidermide di contribuire, assieme ai tessuti sottostanti, alla colorazione delle foglie,fiori, fusto.
Tra i pigmenti sono abondanti:
- Antociani;
- Flavoni;
- Xantoni
I plastidi sono scarsi . Sono presenti cromoplasti in certi fiori e cloroplasti in piante che si trovano in zone scarsamente illuminate, tipo felci. Nella parte area le funzioni tegumentali sono facilitate dall'ispessimento della parete cellulare.
In ambienti umidi c'è un minore ispessimento della parete a causa di una minore perdita di acqua.In ambienti aridi succede il contrario.
La MINERALZZAZIONE DELLA PARETE, è un processo che le cellule attuano per aumentare la resistenza esterna dell'epidermide.
Un'altra e non frequente modificazione della parete è la LIGNIFICAZIONE,che interessa principalmente gli strati piu esterni della parete(felci, foglie di conifera, alloro, ecc.), in altre cellule epidermiche avviene il fenomeno della GELITIFICAZIONE (malvacee e rutacee) nei semi di alcune piante, Lino, tutta l'epidermide ha la parete gelificate. Una delle caratteristiche piu importanti delle pareti delle cellule epidermiche è la presenza della cutina, che impregna le pareti stesse (CUTINIZZAZIONE) e costituisce uno strato superficiale di cutina.
La cutina per la sua impermeabilità all'acqua e ai gas espleta una funzione di protezione efficace contra la perdita di acqua e gas,e difesa contro i parassiti.
Lo spessore della cuticola varia a secondo se si parla di piante xerifile o acquatiche. Nelle piante desertiche ovviamente la cuticola è più spessa.
.
STRUTTURA CUTICOLA-----> SOSTAZA IDROFOBA----->POLIMERO COSTITUITO DA ACIDI GRASSI A LUNGA CATENA (C16-C18) ESTERIFICATI TRA LORO A FORMARE UN RETICOLO TRIDIMENSIONALE.
MINERALIZZAZIONE--------> (SiO2) SILICIZZAZIONE FOGLIE GRAMINACEE, PALME, EQUISETI. CaCO3 CALCIFICAZIONE.
I tricomi dell'ortica presentano entrambi i fenomeni.
Altre strutture dell'epidermide
Tricomi (o peli radicali), generalmente sono costituiti da una singola cellula oppure da complessi di cellule.
Stomi (stoma=bocca) struttura annessa al tessuto epidermico con fuzioni di scambi gassosi,traspirazione.
Dizionario.
Esterificazione: è una reazione chimica tra un alcool e un acido. Spesso l'acido è l'acido carbossilico (-COOH) ma l'esterificazione può avvenire anche con acidi inorganici (A. Fosforico,Solforico,ecc.)
STOMA -----> dal greco bocca

Esoderma, è un tessuto tegumentale primario esterno che si forma per la modificazione secondaria (suberificazione) delle pareti cellulari in tessuto adulto primario, quasi sempre un tessuto parenchimatico privo di spazzi intercellulari.
Suberina------>sostanza idrofoba-------->poliesteri di acidi organici come l'acido suberinico e fellonico uniti a composti aromatici.
L'ESODERMA è localizzata nelle radici sebene possa comparite anche nei fusti sotteranei. Esso può essere mono o pluristratificato.
L'ESODERMA SI SVILUPPA DOPO LA MORTE DEL RIZODERMA
TESSUTI TEGUMENTALI INTERNI
ENDODERMA,è un tessuto tipico degli organi che svolgono la funzione assorbente ed è caratteristico delle radici.
l'endoderma è costituito da un monostrato di cellule vive e prive di spazi intercellulari.
ENDODERMA---- >RADICI------->FUNZIONE ASSORBENTE------->CELLULE VIVE----> PRIVE DI SPAZI INTERCELLULARI --------> IMPEDIRE LIBERO ACCESSO SOSTANZE DANNOSE AI TESSUTI CONDUTTORI.
Apoplasto
Per le proprietà idrofile dei suoi componenti (cellulosa, emicellulose, pectine) la parete cellulare è permeabile all'acqua: l'acqua può agevolmente fluire attraverso le pareti cellulari senza che debba necessariamente attraversare membrane ed entrare quindi nel protoplasto. L'apoplasto è pertanto costituito dall'insieme delle pareti che non sono state rese impermeabili all'acqua a causa di modificazioni con sostanze idrofobe (es.: lignificazione, cutinizzazione, suberificazione). L'apoplasto comprende anche il lume degli elementi vascolari, entro cui l'acqua può liberamente fluire con la corrente traspiratoria.
Simplasto
L'insieme dei protoplasti comunicanti tra loro è detto simplasto. Il simplasto è dato dalle cellule adiacenti il cui citoplasma (ovvero la matrice liquida in esse contenuta) è in comunicazione attraverso dei ponti (nello specifico mettono in comunicazione il reticolo endoplasmatico di due cellule adiacenti) chiamati plasmodesmi. L'acqua che è trasportata per via simplastica non può fluire liberamente come nell'apoplasto; segue due vie: quella attraverso la membrana o attraverso i plasmodesmi. Il simplasto rappresenta un rigoroso sistema di regolazione del flusso di acqua e delle sostanze che la pianta assorbe attraverso le radici. Inoltre in altri distretti della pianta, la presenza di domini simplastici individua insiemi di cellule che sono in relazione tra loro per gli scambi di sostanze, come regolatori di crescita, ioni, zuccheri. Esiste una certa plasticità dei domini simplastici, ottenuta variando il numero di plasmodesmi che collegano le cellule, agevolando i trasporti in determinate direzioni, ostacolandoli in altre. Ad esempio, le cellule degli stomi sono isolate dal simplasto delle cellule epidermiche della foglia. Oppure, in alcuni casi è la via simplastica ad essere utilizzata per il caricamento del floema con zuccheri a livello della foglia.


Approfondimento plasmodesmi
I plasmodesmi, sono strette strisce di citoplasma che connettono i protoplasti di cellule vegetali vicine, sono vie importanti per la comunicazione da cellula a cellula. In quanto stettamente connesso dai plasmodesmi, l'insieme di tutti i protoplasti del corpo della pianta e dei loro plasmodesmi costituisce un continum chiamato simplasto. Il movimento delle sostanze da cellula a cellula tramite i plasmodesmi è detto trasporto smplastico.
I plasmodesmi possono essere classificati in :
- plasmodesmi primari se si formano durante la citocinesi;
- Plasmodesmi secondari se si formano nella parete cellulare de novo.
Come si osserva dal microscopio elettronico, il plasmodesma appare come un canale allineato alla membrana plasmatica e attraversato da una briglia tubolare di reticolo endoplasmatico strettamente ristretto, chiamato DESMOTUBOLO, che è in continuità con ER della cellula adiacente.
I plasmodesmi sono strutture dinamiche in grado di controllare, a vari gradi, il movimento intercellulare delle piccole molecole, e che hanno la capacità di mediare il trasporto di macromolecole, come proteine e un ampio spettro di RNA svolgendo cosi un ruolo prmario nella coordinazione della crescita e dello sviluppo della pianta.
TRASPORTO ACQUA NELLA PIANTA
VIA APOPLASTICA---------> CELLULE ADIACENTI ATTRAVERSO LA PARETE CELLULUARE NON MODIFICATA.
VIA SINPLASTICA-------> ATTRAVERSO ------>PLASMODESMI E PLASMALEMMA
BANDA DEL CASPARY (SCOPERTA 1865) è costituita uno spesso strato di materia idrofoba interposto tra le cellule che blocca il passaggio per via apoplastica verso il cilindro centrale della radice.
Se si osserva una sezione trasversale di molte radici, si osserva che la corteccia occupa l'area più ampia del corpo primario. i plastidi delle cellule corticali comunemente immagazzinano amido e sono generalmente prive di clorofilla.
Le radici delle gimnosperme e angiosperme, che vanno incontro ad una crescita secondaria rilevante, perdono precocemente la loro corteccia. In queste radici le loro cellule corticali rimangono parenchimatiche., al contrario in molte monocotiledoni e nelle eucotiledoni erbacee formate interamente da tessuti primari, la cortecci si conserva per tutta la vita della radice e molte cellule corticali sviluppano pareti secondarie che diventano lignificate.
Indipendentemente dal grado di differenziamento, il tessuto corticale contiene numerosi spazzi intercellulari, spazzi aeriferi necessari per l'areazione delle cellule della radice.
Le cellule corticali hanno numerosi punti in contatto tra loro e i loro protoplasti sono interconessi con i plasmodesmi. Percio le sostanze che si muovono attraverso la corteccia possono seguire la via simplastica (trasporto simplastico) muovendosi tra un protoplasto all'altro mediante i plasmodesmi, o la via apoplastica attraverso gli spazi intercellulari e le maglie delle pareti cellulari o entrambe.
a differenza del resto della corteccia, lo strato più interno è disposto in modo compatto ed è privo di spazi aeriferi. Questo strado è detto endoderma, è caratterizzato dalla presenza delle bande del Caspary nelle sue pareti anticlinali( le pareti radiali e trasversali che sono perpendicolari alla superficie della radice).
La Banda del Caspary non è semplicemente un ispessimento della parete cellulare, ma una porzione integrante di parete primaria e lamella mediana( cioè lo strato di materiale intercellulare che congiunge celleule adiacenti) che è impregnato di suberina e talvolta anche di lignina. La suberina e la lignina infiltrano gli spazi della parete solitamente occupati dall'acqua, conferendo proprietà idrofobe a queste specifiche regioni della parete cellulare.
Le membrane plasmatiche delle cellule endodermiche sono attaccate piuttosto fermamente dalle bande del Caspary. poichè l'endoderma è compatto e le bande del Caspary sono impermeabili all'acqua e agli ioni, il movimento apoplastico dell'acqua e dei soluti attraverso l'endoderma viene bloccato da tali bande.
Quindi , tutte le sostanze che entrano e/o lasciano il cilindro vascolare devono passare attraverso il protoplasto delle cellule endotermiche. Questo passaggio si compie sia attraverso la menbrana del protoplasta o attraverso i plasodesmi che connettono le cellule dell'endodermiche con i protoplasti delle cellule vicine sia della corteccia che del cilindo vascolare.
Se si osserva una sezione trasversale di molte radici, si osserva che la corteccia occupa l'area più ampia del corpo primario. i plastidi delle cellule corticali comunemente immagazzinano amido e sono generalmente prive di clorofilla.
Le radici delle gimnosperme e angiosperme, che vanno incontro ad una crescita secondaria rilevante, perdono precocemente la loro corteccia. In queste radici le loro cellule corticali rimangono parenchimatiche., al contrario in molte monocotiledoni e nelle eucotiledoni erbacee formate interamente da tessuti primari, la cortecci si conserva per tutta la vita della radice e molte cellule corticali sviluppano pareti secondarie che diventano lignificate.
Indipendentemente dal grado di differenziamento, il tessuto corticale contiene numerosi spazzi intercellulari, spazzi aeriferi necessari per l'areazione delle cellule della radice.
Le cellule corticali hanno numerosi punti in contatto tra loro e i loro protoplasti sono interconessi con i plasmodesmi. Percio le sostanze che si muovono attraverso la corteccia possono seguire la via simplastica (trasporto simplastico) muovendosi tra un protoplasto all'altro mediante i plasmodesmi, o la via apoplastica attraverso gli spazi intercellulari e le maglie delle pareti cellulari o entrambe.
a differenza del resto della corteccia, lo strato più interno è disposto in modo compatto ed è privo di spazi aeriferi. Questo strado è detto endoderma, è caratterizzato dalla presenza delle bande del Caspary nelle sue pareti anticlinali( le pareti radiali e trasversali che sono perpendicolari alla superficie della radice).
La Banda del Caspary non è semplicemente un ispessimento della parete cellulare, ma una porzione integrante di parete primaria e lamella mediana( cioè lo strato di materiale intercellulare che congiunge celleule adiacenti) che è impregnato di suberina e talvolta anche di lignina. La suberina e la lignina infiltrano gli spazi della parete solitamente occupati dall'acqua, conferendo proprietà idrofobe a queste specifiche regioni della parete cellulare.
Le membrane plasmatiche delle cellule endodermiche sono attaccate piuttosto fermamente dalle bande del Caspary. poichè l'endoderma è compatto e le bande del Caspary sono impermeabili all'acqua e agli ioni, il movimento apoplastico dell'acqua e dei soluti attraverso l'endoderma viene bloccato da tali bande.
Quindi , tutte le sostanze che entrano e/o lasciano il cilindro vascolare devono passare attraverso il protoplasto delle cellule endotermiche. Questo passaggio si compie sia attraverso la menbrana del protoplasta o attraverso i plasodesmi che connettono le cellule dell'endodermiche con i protoplasti delle cellule vicine sia della corteccia che del cilindo vascolare.
TESSUTI TEGUMENTALI SECONDARI: SUGHERO
L'accrescimento secondario provocato dal cambio cribo-legnoso, con la produzione di nuovo legno e nuovo libro, determina un ingrossamento del corpo secondario della pianta. Di conseguenza gli strati più esterni, in particolare l'epidermide, non sono in grado di seguire tale accrescimento se non per un tempo limitato, pertanto l'epidermide viene sostituita dal peri-derma.
Il SUGHERO è un tessuto tegumentale secondario esterno, la cui funzione è quella di sostituire con efficacia maggiore, l'epidermide, quando in seguito all'accrescimento secondario questo non è pù efficace nel ricoprire l'intero corpo della pianta e pertanto viene lacerata ed eliminata.
A differenza dell'epidermide il sughero è formato da cellule morte e costituiscono il tipico tessuto tegumentale secondario della pianta
Oltre che sul fusto e sulle radici, il tessuto suberoso si trova nei tuberi (esempio patata) nei frutti succulenti (pere, mele, soprattutto nelle varietà invernali) e nelle squame delle gemme.
Inoltre il sughero puo essere formato come tessuto cicatriziale per riparare ferite.
Il sughero è privo di spazzi intercellulari e formato di cellule prismatiche, disposta in fila l'una sopra all'atra.
Le cellule possono avere le pareti ispessite e suberificate su tutti i lati oppure su uno soltatnto.
Il processo di suberificazione avviene con l'apposizione di lamelle di suberina a ridosso della parete primaria; molto spesso altri strati cellulosici opposti contra la lamella di suberina, formando la parete terziaria, che può lignificare..
Alla fine dell'opposizione le cellule muoiono, si riempiono di aria oppure di materiale di colore scuro, costituito da resine o da derivati tannici (flobafeni), a cui il sughero deve in gran parte la sua colorazione.
Per le proprietà delle sue pareti il sughero è praticamente impermeabile all'acqua e ai gas.
Vi sono piante in cui il sughero ha struttura omogenea, altre con struttura stratificata in quanto in esso si alternano strati con pareti sottili e strati con pareti ispessite, come ad esempio il sughero di betulla.
La continuità del sughero viene interotta dalle lenticelle, che sono strutture caratterizzate da cellule arrotondate con pareti poco o per nulla suberificate e che lasciano tra loro parecchi spazi intercellulari attraverso i quali l'aria può arrivare ai tessuti sottostanti.
Le lenticelle sono presenti in numero minore rispetto a quello degli stomi, pur formandosi in corrispondenza di questi. Nel processo di formazione della lenticella, le cellule del parenchima sotto epidermico, limitante la camera sottostomatica, subiscono una seri di divisioni tutte parallele all'epidermide. La clorofilla scompare, mentre le cellule che si dividono si trovano ad essere situate sempre più profondamente, formando un tessuto incolore tessuto di riempimento.
IL MERISTEMA CHE SI FORMA SOTTO OGNI LENTICELLA VIENE CHIAMATO FELLOGENO DELLA LENTICELLA ED E' IN CONTINUITA' CON IL FELLOGENO.
La massa di tessuto di tessuto di riempimento va progressivamente aumentando, fino a lacerare l'epidermide. Le cellule a contatto con l'aria muoio e facilmente si sfalòdano; in loro sostituzione il fellogeno dlla lenticella continua a produrre nuove cellule internamente a preesistente tessuto di riempimento; le cellule di riempimento conservano le pareti sottili e molte volte non suberificate.
Nella stagione inverale le lenticelle vengono chiuse, in quanto il fellogeno delle lenticelle forma un piccolo strato di sughero, con cellule prive di spazi intercellulari e con pareti suberificate.
A termine dell'inverno, il medesimo fellogeno delle lenticelle o una di nuova formazione, forma delle cellule sotto lo strato di chiusura, che premono fino a lacerare tale strato, consentendo la riapertura delle lenticelle.

RIASSUNTO TESSUTI TEGUMENTALI
Il tessuto meristematico secondario si forma dal sdiferenziamento di cellule adulte quasi sempre parenchimatiche che riaquistano la capacità di dividersi per scissione minaria della cellula.
FELLOGENO------>MERISTEMA SECONDARIO TEGUMENTALE------> GENERA IL TESSUTO SECONDARIO TEGUMENTALE IL PERIDERMA (SUGHERO E FELLODERMA)
Il fellogeno detto anche-------> CAMBIO DEL SUGHERO OPPURE CAMBIO-SUBERO-FELLODERMICO
IL FELLOGENO GENERA------>Periderma--------------> Sughero verso l'esterno e felloderma verso l'iterno
SUGHERO---------> CELLULE MORTE
FELLODERMA--------> CELLULE PARENCHIMATICHE VIVE
SCLERENCHIMA
Differisce dal collenchima perchè costituito da cellule dopo aver completato il proprio differenziamento muoipno, in quanto i protoplasti subiscono la morte cellulare programmata, PCD (Programmed Cell Death).
Lo scherenchima è il tessuto tipico delle pareti della painta che hanno completato la fase di differenziamento e crescita per distensione.
Rispetto al Collenchima è molto più resistente, perchè le pareti delle sue cellule vanno incontro modificazioni, con opposizione di strati di parete secondaria omogeneamente distribuiti e lignificati.
In base alla loro morfologia si possono distinguere due titpi di clerenchima:
Le scleridi sono cellule isodiametriche, a volte allungate e brevemente ramificat, si formano in seguito a differenziamento di cellule parenchimatichedi origine primaria e secondaria.
La loro parete è lignificate ed ispissita, solcata da parechi porocanali, presentano forme molto diverse e tendono ad essere sviluppate euguale in tutte le direzioni.
Sono presenti in tutto il corpo della pianta (fusto,foglie, e semi)isolate o riunite in gruppi, come le cellule pietrose della polpa della pera, o in ammassi consistenti, come nel guscio di noci o nocciole.
Le scleridi vengono chiamate con nomi diverse in base alla loro morfologia:
FIBRE
Le fibre sono cellule allungate in senso assiale ed afusolate all'estrmità, in sezione trasversale appaiono poligonali, possono essere isolate o aggregate in fasci o cordoni longitudinali.
La loro parete è ispessita e il lume cellulare è parecchio ridotto.
Esistono due tipi di fibre:
Le fibre xilematiche sono le fibre dello XILEMA, anche dette fibre libriforme o cellule libriforme per la loro somiglianza alle fibre floematiche, si trovano nel legno specialmente secondario delle angiosperme , del quale sono parte integrante insieme ai vasi e alle cellelu parenchimatiche. Sono assai variabili per aspetto, grandezza spessore della parete tipo e freguenza della punteggiatura; hanno però la caratteristica comune di avere la parete secondaria sempre lignificata.delle volte sono settate per la presenz di pareti trasversali, che si formano dopo che,su quelle longitudinali, sono stati depositati gli stradi secondari d'ispessimento.
In questo caso, esse rimangono vive per lungo tempo e possono contenere amido, oli, resine o cristalli di ossalto di calcio.






















Le cellule possono avere le pareti ispessite e suberificate su tutti i lati oppure su uno soltatnto.
Il processo di suberificazione avviene con l'apposizione di lamelle di suberina a ridosso della parete primaria; molto spesso altri strati cellulosici opposti contra la lamella di suberina, formando la parete terziaria, che può lignificare..
Alla fine dell'opposizione le cellule muoiono, si riempiono di aria oppure di materiale di colore scuro, costituito da resine o da derivati tannici (flobafeni), a cui il sughero deve in gran parte la sua colorazione.
Per le proprietà delle sue pareti il sughero è praticamente impermeabile all'acqua e ai gas.
Vi sono piante in cui il sughero ha struttura omogenea, altre con struttura stratificata in quanto in esso si alternano strati con pareti sottili e strati con pareti ispessite, come ad esempio il sughero di betulla.
La continuità del sughero viene interotta dalle lenticelle, che sono strutture caratterizzate da cellule arrotondate con pareti poco o per nulla suberificate e che lasciano tra loro parecchi spazi intercellulari attraverso i quali l'aria può arrivare ai tessuti sottostanti.
Le lenticelle sono presenti in numero minore rispetto a quello degli stomi, pur formandosi in corrispondenza di questi. Nel processo di formazione della lenticella, le cellule del parenchima sotto epidermico, limitante la camera sottostomatica, subiscono una seri di divisioni tutte parallele all'epidermide. La clorofilla scompare, mentre le cellule che si dividono si trovano ad essere situate sempre più profondamente, formando un tessuto incolore tessuto di riempimento.
IL MERISTEMA CHE SI FORMA SOTTO OGNI LENTICELLA VIENE CHIAMATO FELLOGENO DELLA LENTICELLA ED E' IN CONTINUITA' CON IL FELLOGENO.
La massa di tessuto di tessuto di riempimento va progressivamente aumentando, fino a lacerare l'epidermide. Le cellule a contatto con l'aria muoio e facilmente si sfalòdano; in loro sostituzione il fellogeno dlla lenticella continua a produrre nuove cellule internamente a preesistente tessuto di riempimento; le cellule di riempimento conservano le pareti sottili e molte volte non suberificate.
Nella stagione inverale le lenticelle vengono chiuse, in quanto il fellogeno delle lenticelle forma un piccolo strato di sughero, con cellule prive di spazi intercellulari e con pareti suberificate.
A termine dell'inverno, il medesimo fellogeno delle lenticelle o una di nuova formazione, forma delle cellule sotto lo strato di chiusura, che premono fino a lacerare tale strato, consentendo la riapertura delle lenticelle.

RIASSUNTO TESSUTI TEGUMENTALI
Il tessuto meristematico secondario si forma dal sdiferenziamento di cellule adulte quasi sempre parenchimatiche che riaquistano la capacità di dividersi per scissione minaria della cellula.
FELLOGENO------>MERISTEMA SECONDARIO TEGUMENTALE------> GENERA IL TESSUTO SECONDARIO TEGUMENTALE IL PERIDERMA (SUGHERO E FELLODERMA)
Il fellogeno detto anche-------> CAMBIO DEL SUGHERO OPPURE CAMBIO-SUBERO-FELLODERMICO
IL FELLOGENO GENERA------>Periderma--------------> Sughero verso l'esterno e felloderma verso l'iterno
SUGHERO---------> CELLULE MORTE
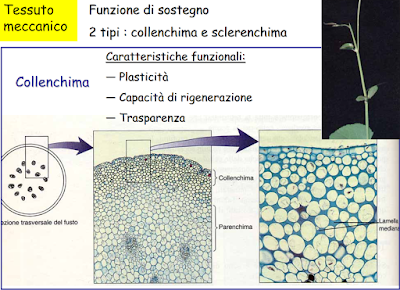
TESSUTI MECCANICI
Le funzioni fondamentali di questi tessuti è quello di sostenere il corpo della pianta e di fornire resistenza al piegamento, sopratutto nelle piante erbacee, e alla trazione. Le proprietà meccaniche di questi tessuti sono da attribuire alla composizione chmica e alla struttura della pareti cellulari che li costituiscono.
Sulla base della natura e delle caratteristiche delle pareti cellulari si distinguono due tipi di tessuti meccanici:
Collenchima e Sclerenchima; il primo è costituito da cellule vive, il secondo da cellule morte.
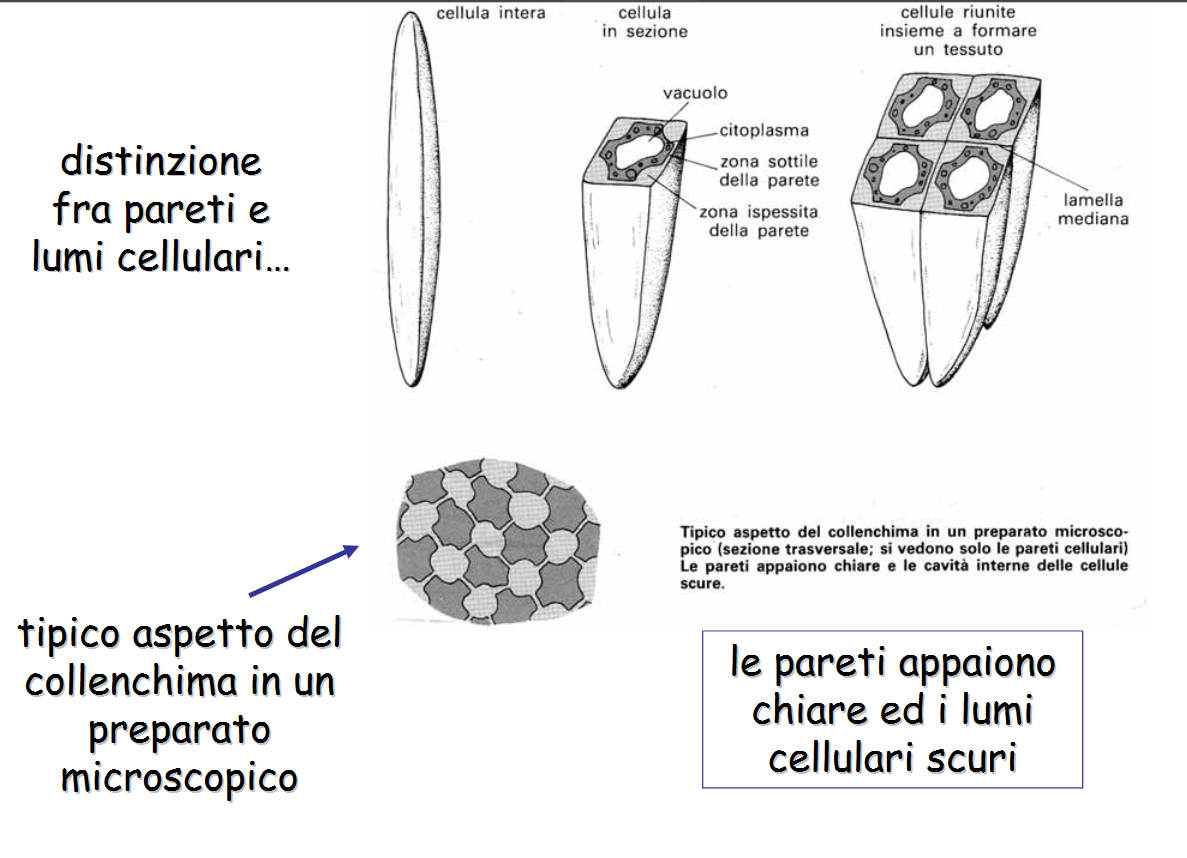
il collenchima è un tessuto adulto primario situato in posizione periferica, sotto l'epidermide o separato dall'epidermide da pochi stradi di cellule parenchimatiche. Il collenchima è più comunemente osservato eudicotiledoni, mentre è assente nei fusti e foglie di molti felci e monocotiledoni (erbaceee, inclusi i cereali) che differenziano presto lo schelernchima. Si trova nelle parti del germoglio che sono ancora in accrescimento ed è costituito da cellule vive, isodiamentriche oppure allungate longitudinalmente, a sezione poligonale, affusolatwe o tronche agli estremi.
La parete è celluloso-pectica, non lignificata, ispessita in determinate aree. Il collenchima come il parenchima ha pareti cellulari in grado di stirarsi e/0 crescere durante il differenziamento.
Alcune volte nelle cellule sono presenti cloroplasti, che le rendono simili alle cellul parenchimatiche, ma l'ispessimento delle pareti le rende riconoscibili come cellule collenchimatiche.
La parete delle celleule parenchimatiche è una parete primaria formata interamente dallo stesso numero di strati, caratterizzati in alcuni punti da diverso spessore.
Talli ispessimenti cominciano a comparire prima che sia terminato l'accrescimento per distensione delle cellule stesse.
Sulla base dei punti ispessiti possiamo distinguere diversi tipi di collenchima:
- Collenchima angolare, se gli ispessimenti sono gli ngoli dove confluiscono più cellule;
- Collenchima Lamellare se gli ispessimenti riguardano le pareti tangenziali;
- Collenchima anulare se le pareti sono uniformemente ispessite e il lume cellulare è pressapoco circolare.
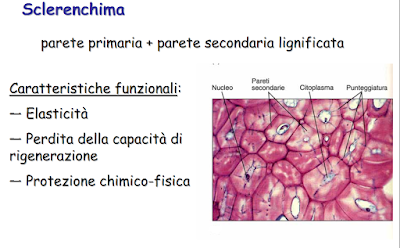
SCLERENCHIMA
Differisce dal collenchima perchè costituito da cellule dopo aver completato il proprio differenziamento muoipno, in quanto i protoplasti subiscono la morte cellulare programmata, PCD (Programmed Cell Death).
Lo scherenchima è il tessuto tipico delle pareti della painta che hanno completato la fase di differenziamento e crescita per distensione.
Rispetto al Collenchima è molto più resistente, perchè le pareti delle sue cellule vanno incontro modificazioni, con opposizione di strati di parete secondaria omogeneamente distribuiti e lignificati.
In base alla loro morfologia si possono distinguere due titpi di clerenchima:
- Scleridi;
- Fibre.
Le scleridi sono cellule isodiametriche, a volte allungate e brevemente ramificat, si formano in seguito a differenziamento di cellule parenchimatichedi origine primaria e secondaria.
La loro parete è lignificate ed ispissita, solcata da parechi porocanali, presentano forme molto diverse e tendono ad essere sviluppate euguale in tutte le direzioni.
Sono presenti in tutto il corpo della pianta (fusto,foglie, e semi)isolate o riunite in gruppi, come le cellule pietrose della polpa della pera, o in ammassi consistenti, come nel guscio di noci o nocciole.
Le scleridi vengono chiamate con nomi diverse in base alla loro morfologia:
- astrosclereridi (cellulle ramificate o più o meno a forma di stella, come nella foglia di the);
- brachiscleridi (tondeggianti o poligonali, come nella polpa di pera);
- macroscleridi (a forma di bastoncino corto, come quelle del guscio del seme di fagiolo, piselli ed altri legumi);
- osteoscleridi (a forma di osso, come quelle che si trovano nelle foglie di molte dicotiledoni);
- tricoscleridi ( a forma di capello, come lescleridi delle foglie di olivo).
FIBRE
Le fibre sono cellule allungate in senso assiale ed afusolate all'estrmità, in sezione trasversale appaiono poligonali, possono essere isolate o aggregate in fasci o cordoni longitudinali.
La loro parete è ispessita e il lume cellulare è parecchio ridotto.
Esistono due tipi di fibre:
- fibrexilemetiche, o fibre xilari;
- fibre extraxilari;
Le fibre xilematiche sono le fibre dello XILEMA, anche dette fibre libriforme o cellule libriforme per la loro somiglianza alle fibre floematiche, si trovano nel legno specialmente secondario delle angiosperme , del quale sono parte integrante insieme ai vasi e alle cellelu parenchimatiche. Sono assai variabili per aspetto, grandezza spessore della parete tipo e freguenza della punteggiatura; hanno però la caratteristica comune di avere la parete secondaria sempre lignificata.delle volte sono settate per la presenz di pareti trasversali, che si formano dopo che,su quelle longitudinali, sono stati depositati gli stradi secondari d'ispessimento.
In questo caso, esse rimangono vive per lungo tempo e possono contenere amido, oli, resine o cristalli di ossalto di calcio.























TESSUTI CONDUTTORI
Salve Dottor Coccia, la ringrazio per questi fantastici lavori.Ottima spiegazione e sopratutto organizzazione.Volevo chiederle se fosse possibile ricaricare la parte sui tessuti conduttori che sembra essere cancellata. Le auguro tante buone ricerche. Buon lavoro.
RispondiElimina